Карта слов и выражений русского языка
Онлайн-тезаурус с возможностью поиска ассоциаций, синонимов, контекстных связей
и
примеров
предложений к словам и выражениям русского языка.
Справочная информация по склонению имён существительных и прилагательных,
спряжению
глаголов, а также
морфемному строению слов.
Сайт оснащён мощной системой поиска с
поддержкой русской морфологии.
Разбор слова
по составу ОНЛАЙН
Подобрать синонимы
ОНЛАЙН
Найти предложения со словом
или
выражением ОНЛАЙН
Поиск по произведениям русской классики
ОНЛАЙН
Словарь афоризмов русских писателей
На букву М Со слова «мембранный»
Фраза «мембранный потенциал»
Фраза состоит из двух слов и 19 букв без пробелов.
- Синонимы к фразе
- Написание фразы наоборот
- Написание фразы в транслите
- Написание фразы шрифтом Брайля
- Передача фразы на азбуке Морзе
- Произношение фразы на дактильной азбуке
- Остальные фразы со слова «мембранный»
- Остальные фразы из 2 слов
 07:54
07:54
Физиология. Потенциал действия
 08:26
08:26
Мембранные потенциалы — Часть 1
 06:49
06:49
Потенциал покоя: — 70 мВ. Деполяризация, реполяризация
 05:03
05:03
Мембранный потенциал
 08:50
08:50
Мембранные потенциалы — Часть 2
 13:31
13:31
Происхождение потенциала покоя
Синонимы к фразе «мембранный потенциал»
Какие близкие по смыслу слова и фразы, а также похожие выражения существуют. Как можно написать по-другому или сказать другими словами.
Фразы
- + адекватный раздражитель −
- + азотистый баланс −
- + биохимические реакции −
- + биоэлектрический потенциал −
- + водитель ритма −
- + вынужденные колебания −
- + градиент концентрации −
- + дыхательная цепь −
- + избирательная проницаемость −
- + калиевые каналы −
- + кислородная ёмкость крови −
- + лёгочные капилляры −
- + мембранный потенциал −
- + метаболические процессы −
- + мышечные клетки −
- + обратное всасывание −
- + окислительное фосфорилирование −
- + пассивный транспорт −
- + первичная моча −
- + перенос электрона −
- + положительные ионы −
- + потенциал действия −
- + потенциал покоя −
- + промежуточные филаменты −
Ваш синоним добавлен!
Написание фразы «мембранный потенциал» наоборот
Как эта фраза пишется в обратной последовательности.
лаицнетоп йыннарбмем 😀
Написание фразы «мембранный потенциал» в транслите
Как эта фраза пишется в транслитерации.
в армянской🇦🇲 մեմբրաննըյ պոտենցիալ
в латинской🇬🇧 membranny potentsial
Как эта фраза пишется в пьюникоде — Punycode, ACE-последовательность IDN
xn--80ablslbjay7i xn--80ajjnhegz2b
Как эта фраза пишется в английской Qwerty-раскладке клавиатуры.
vtv,hfyysqgjntywbfk
Написание фразы «мембранный потенциал» шрифтом Брайля
Как эта фраза пишется рельефно-точечным тактильным шрифтом.
⠍⠑⠍⠃⠗⠁⠝⠝⠮⠯⠀⠏⠕⠞⠑⠝⠉⠊⠁⠇
Передача фразы «мембранный потенциал» на азбуке Морзе
Как эта фраза передаётся на морзянке.
– – ⋅ – – – ⋅ ⋅ ⋅ ⋅ – ⋅ ⋅ – – ⋅ – ⋅ – ⋅ – – ⋅ – – – ⋅ – – ⋅ – – – – ⋅ – ⋅ – ⋅ – ⋅ ⋅ ⋅ ⋅ – ⋅ – ⋅ ⋅
Произношение фразы «мембранный потенциал» на дактильной азбуке
Как эта фраза произносится на ручной азбуке глухонемых (но не на языке жестов).
Передача фразы «мембранный потенциал» семафорной азбукой
Как эта фраза передаётся флажковой сигнализацией.













Остальные фразы со слова «мембранный»
Какие ещё фразы начинаются с этого слова.
- мембранный гидроаккумулятор
- мембранный насос
- мембранный транспорт
Ваша фраза добавлена!
Остальные фразы из 2 слов
Какие ещё фразы состоят из такого же количества слов.
- а вдобавок
- а вдруг
- а ведь
- а вот
- а если
- а ещё
- а именно
- а капелла
- а каторга
- а ну-ка
- а приятно
- а также
- а там
- а то
- аа говорит
- аа отвечает
- аа рассказывает
- ааронов жезл
- аароново благословение
- аароново согласие
- аб ово
- абажур лампы
- абазинская аристократия
- абазинская литература
Комментарии
 @tobx 05.01.2020
@tobx 05.01.2020 02:24
Что значит фраза «мембранный потенциал»? Как это понять?..
Ответить
@jlopveo 07.09.2022 06:05
![]() 1
1
×
![]()
Здравствуйте!
У вас есть вопрос или вам нужна помощь?
![]()
Спасибо, ваш вопрос принят.
Ответ на него появится на сайте в ближайшее время.
А Б В Г Д Е Ё Ж З И Й К Л М Н О П Р С Т У Ф Х Ц Ч Ш Щ Ъ Ы Ь Э Ю Я
Транслит Пьюникод Шрифт Брайля Азбука Морзе Дактильная азбука Семафорная азбука
Палиндромы Сантана
Народный словарь великого и могучего живого великорусского языка.
Онлайн-словарь слов и выражений русского языка. Ассоциации к словам, синонимы слов, сочетаемость фраз. Морфологический разбор: склонение существительных и прилагательных, а также спряжение глаголов. Морфемный разбор по составу словоформ.
По всем вопросам просьба обращаться в письмошную.
1
Первый слайд презентации: Возбудимые ткани. Биоэлектрические явления в возбудимых тканях и методы их исследования
Лекция № 3 (первая часть)
9.09.08

Изображение слайда
История изучения биоэлектрических явлений (опыты Л.Гальвани)

Изображение слайда
3
Слайд 3: История изучения биоэлектрических явлений
Первые данные о существовании биоэлектрических явлений («животного электричества») были получены в третьей четверти XVIII в. при изучении природы электрического разряда, наносимого некоторыми p ыбами при защите и нападении.

Изображение слайда
4
Слайд 4: История изучения биоэлектрических явлений
Начало систематического изучения биоэлектрических явлений связывают с именем итальянского физика и анатома Луиджи Гальвани ( Galvani L.).
Л.Гальвани первым убедился в существовании «живого электричества». Термин «животное электричество» принадлежит Л.Гальвани.
Это произошло в 1771 г. По данным других источников — в 1780 г., но опубликованы результаты открытия были только в 1791 г.

Изображение слайда
5
Слайд 5: Луи́джи Гальва́ни (Luigi Galvani, 1737 — 1798 )
— итальянский врач, анатом, физиолог и физик, один из основателей электрофизиологии, основоположник экспериментальной электрофизиологии. Первым исследовал электрические явления при мышечном сокращении («животное электричество»).

Изображение слайда
6
Слайд 6: Сочинения Гальва́ни
Трактат о силах электричества при мышечном движении (De Viribus Electricatitis in Motu Musculari Commentarius) (1791)

Изображение слайда
7
Слайд 7: Гальвани с женой и помощником проводят эксперимент в домашней лаборатории. А. Муцци, 1862 год

Изображение слайда
8
Слайд 8: Лаборатория Гальвани

Изображение слайда
9
Слайд 9: Схема опыта по изучению атмосферного электричества
Детектором служит лягушачья лапка, нерв которой соединен с громоотводом, а мышца соединена через проводник с водой в колодце. Рисунок из трактата Гальвани.

Изображение слайда
10
Слайд 10: История изучения биоэлектрических явлений
Первый («балконный») опыт Л.Гальвани
Учёного интересовало влияние электрических грозовых разрядов на мышцы лягушки.
Препарат задних лапок лягушек на медном крючке был подвешен в грозу к железному балкону.
Влияние молнии на мышцы лягушки он не заметил, но отметил другое — от ветра в дождь препарат задевал балконные перила, и в этот момент мышцы сокращались.

Изображение слайда
11
Слайд 11: История изучения биоэлектрических явлений
Первый («балконный») опыт Л.Гальвани

Изображение слайда
12
Слайд 12: История изучения биоэлектрических явлений
Первый («балконный») опыт Л.Гальвани

Изображение слайда
Первый («балконный») опыт Л.Гальвани

Изображение слайда
14
Слайд 14: История изучения биоэлектрических явлений
Л.Гальвани считал, что мышцы и нервы заряжены электричеством («живое электричество’‘) наподобие лейденской банки (конденсатора). Замыкания цепи тока вызывает сокращение.

Изображение слайда
15
Слайд 15: Пара лейденских банок, изготовленная студентами в 1890–1910 гг

Изображение слайда
16
Слайд 16: История изучения биоэлектрических явлений
А. Вольта ( Volta A.) опроверг это объяснение и доказал, что источником электрического тока является «гальваническая пара» — железо-медь.

Изображение слайда
Вольтов столб, состоящий из металлических дисков, разделенных кружками мокрой ткани.

Изображение слайда
Вольта демонстрирует перед Наполеоном свое изобретение — Вольтов столб. Художник Дж. Бертини. 1801 год.

Изображение слайда
19
Слайд 19: История изучения биоэлектрических явлений
Второй опыт Л.Гальвани
(без металла)
Л.Гальвани поставил в ответ на возражения А.Вольта
Набрасывался нерв между поврежденной (срезом) и неповрежденной поверхностями мышцы
В ответ — сокращение интактной мышцы

Изображение слайда
20
Слайд 20: Второй опыт Л.Гальвани (без металла)

Изображение слайда
21
Слайд 21: История изучения биоэлектрических явлений
Многолетний научный спор (1791-1797) между Л.Гальвани и А.Вольта завершился двумя крупными открытиями:
были установлены факты, свидетельствующие о наличии электрических потенциалов в нервной и мышечной тканях, и
открыт новый способ получения электрического тока при помощи разнородных металлов — co здан гальванический элемент («вольтов столб», 1800).

Изображение слайда
22
Слайд 22: История изучения биоэлектрических явлений
Систематическое исследование потенциалов в мышцах и нервах в состоянии покоя и возбуждения было начато немецким физиологом
Эмилем Дюбуа-Реймоном
( du Bois — Reymond E. ) в 1848 г.
Прямые измерения потенциалов в живых тканях стали возможны только после изобретения гальванометров

Изображение слайда
23
Слайд 23: История изучения биоэлектрических явлений
40—50-е годы XX века:
с помощью внутриклеточных микроэлектродов удалось произвести прямую регистрацию электрических потенциалов клеточных мембран.

Изображение слайда
Понятие «мембранный потенциал»

Изображение слайда
25
Слайд 25: Понятие «мембранный потенциал»
Биологическую мембрану можно рассматривать как электрический конденсатор.

Изображение слайда
26
Слайд 26: Понятие «мембранный потенциал»
Биологическую мембрану можно рассматривать как электрический конденсатор.
Пластинами являются электролиты наружного и внутреннего растворов (внеклеточного и цитоплазмы) с погруженными в них «головами» липидных молекул.
Проводники разделены диэлектрическим слоем, образованным неполярной частью липидных молекул — двойным слоем их «хвостов»

Изображение слайда
27
Слайд 27: Понятие «мембранный потенциал»
Мембранный потенциал – это разность потенциалов между цитоплазмой и окружающим клетку наружным раствором.

Изображение слайда
28
Слайд 28: Понятие «мембранный потенциал»
При измерении мембранного потенциала активный электрод располагают внутри клетки, пассивный – снаружи.

Изображение слайда
29
Слайд 29: Понятие «мембранный потенциал»
МП = ?

Изображение слайда
30
Слайд 30: Понятие «мембранный потенциал»
В электрофизиологии принято задавать потенциал окружающей клетку среды (наружной поверхности мембраны) и придавать ему значение «0 мВ».

Изображение слайда
Мембранный потенциал покоя

Изображение слайда
32
Слайд 32: Мембранный потенциал покоя
Мембранный потенциал покоя (ПП)
— мембранный потенциал клетки в состоянии физиологического покоя

Изображение слайда
33
Слайд 33: Мембранный потенциал покоя
У различных клеток мембранный потенциал покоя варьирует
от –50 мВ
до –90 мВ

Изображение слайда
34
Слайд 34: Мембранный потенциал покоя
Довольно часто термин «мембранный потенциал» используют как синоним термина «потенциал покоя».
На наш взгляд, это недопустимо !!!
Термин «мембранный потенциал» просто необходим для обозначения любого значения трансмембранной разницы потенциала, наблюдаемое как в состоянии покоя клетки, так и при возбуждении, в любое время и в любом состоянии.

Изображение слайда
Регистрация потенциала покоя

Изображение слайда
36
Слайд 36: Регистрация потенциала покоя
методом повреждения
методом внутриклеточного отведения

Изображение слайда
37
Слайд 37: Регистрация потенциала покоя
Схема регистрации мембранного потенциала покоя методом повреждения на макропрепарате.

Изображение слайда
38
Слайд 38: Регистрация потенциала покоя
Схема измерения потенциала покоя клетки с помощью внутриклеточного электрода.

Изображение слайда
39
Слайд 39: Регистрация потенциала покоя
Внутриклеточный метод регистрации ПП
Микроэлектрод устанавливают над исследуемым объектом, например скелетной мышцей, а затем при помощи микроманипулятора вводят внутрь клетки.
При удачном введении микроэлектрода мембрана плотно охватывает его кончик и клетка сохраняет способность функционировать в течение нескольких часов, не проявляя признаков повреждения.
Микроэлектрод является активным (референтным). Электрод сравнения (индифферентный) обычных размеров погружают в нормальный солевой раствор, в котором находится исследуемая ткань.

Изображение слайда
40
Слайд 40: Регистрация потенциала покоя
Результаты измерения разности потенциала микроэлектродным методом при разном расположении активного электрода

Изображение слайда
Механизм формирования (электрогенез) потенциала покоя

Изображение слайда
42
Слайд 42: Механизм формирования потенциала покоя
Значение потенциала покоя клетки определяется двумя основными факторами:
соотношением концентраций проникающих через покоящуюся поверхностную мембрану катионов и анионов
соотношением проницаемостей мембраны для этих ионов
Не говорите величина потенциала покоя !

Изображение слайда
43
Слайд 43: Механизм формирования потенциала покоя
Для количественного описания этой закономерности используют уравнение Гольдмана — Ходжкина — Kатца:
где E m — потенциал покоя; R – газовая постоянная; T – абсолютная температура; F – постоянная Фарадея; Р K, РNa, Р Cl — проницаемости мембраны для ионов K+, Na + и Сl- соответственно; K o +, Na o + и Сl o — — наружные концентрации ионов K +, Na + и Сl -, а K i +, Na i + и Сl i — — их внутренние концентрации.

Изображение слайда
44
Слайд 44: Механизм формирования потенциала покоя
Основным механизмом формирования потенциала покоя являются
создание концентрационной асимметрии K+ при работе калий-натриевого насоса (калий-натриевой АТФазы)
выход K+ из клетки по градиенту концентрации

Изображение слайда

Изображение слайда
Изменения потенциала покоя

Изображение слайда

Изображение слайда
48
Слайд 48: Изменения потенциала покоя

Изображение слайда

Изображение слайда
50
Слайд 50: Изменения потенциала покоя
Уменьшение ПП — деполяризация
(ПП становится менее отрицательным)
Увеличение ПП — гиперполяризация
(ПП становится более отрицательным)

Изображение слайда
Локальный ответ

Изображение слайда

Изображение слайда

Изображение слайда

Изображение слайда
Опыт К.Маттеуччи (вторичный тетанус)

Изображение слайда
56
Слайд 56: Значение слова тетанус
Т е танус — (от греч. tetanos — оцепенение, судорога), состояние длительного сокращения, непрерывного напряжения мышцы

Изображение слайда
57
Слайд 57: Значение слова тетанус
Тетанус возникающее при поступлении к мышце нервных импульсов с такой частотой (более 20 Гц), что расслабления между последовательными одиночными сокращениями не происходит.

Изображение слайда
58
Слайд 58: Значение слова тетанус
Столбняк (тетанус, tetanus, генерализованный, острый, распространенный столбняк) — острое инфекционное заболевание, обусловленное воздействием на организм экзотоксина столбнячной палочки с преимущественным поражением нервной системы, характеризующееся тоническими и судорожными сокращениями поперечно-полосатых мышц.

Изображение слайда

Изображение слайда
60
Слайд 60: Вторичный тетанус

Изображение слайда
61
Слайд 61: Вопрос 9 + 10
Потенциал действия
+
Возбудимость при возбуждении

Изображение слайда
Изображение ПП и ПД в качестве эмблемы Отдела Биофизики Мембран Биофизического Сообщества на медали имени Коула.

Изображение слайда

Изображение слайда
64
Слайд 64: Потенциал действия (физиологический),
быстрое колебание мембранного потенциала, возникающее при возбуждении нервных и мышечных клеток (волокон); активный электрический сигнал, с помощью которого осуществляется передача информации в организме человека и животных. Основан на быстро обратимых изменениях ионной проницаемости клеточной мембраны, связанных с активацией и инактивацией ионных мембранных каналов.

Изображение слайда
65
Слайд 65: Потенциал действия (физиологический)
колебание мембранного потенциала, возникающее при возбуждении, основанное на обратимых изменениях ионной проницаемости клеточной мембраны, связанных с активацией и инактивацией ионных мембранных каналов.

Изображение слайда

Изображение слайда

Изображение слайда
Типы ПД

Изображение слайда

Изображение слайда
Ионный механизм формирования пикообразного ПД

Изображение слайда

Изображение слайда
72
Слайд 72: Раздражение и возбуждение как основные типы реакции тканей на раздражение Подробнее – Учебник том I, С.2 7 (осторожно)
Вопрос 13

Изображение слайда
— неспецифический ответ ткани на действие раздражителя (изменение метаболизма, гиперплазия, гипертрофия …)

Изображение слайда
— специфический электрический ответ ткани на действие раздражителя (генерация потенциала действия, рецепторного потенциала, постсинаптического потенциала …)

Изображение слайда
Ткань изменила метаболизм под действием электрического тока. Это раздражение !!!
При растяжении в ткани возник и распространяется потенциал действия. Это возбуждение.

Изображение слайда
76
Слайд 76: Понятия «ткани», «возбудимые ткани» Подробнее – Учебник том I, С.2 7
Вопрос 14

Изображение слайда
77
Слайд 77: Определение понятия «ткань»
— совокупность гистологических элементов (клеточных и неклеточных), имеющих общность происхождения, строения и функции

Изображение слайда
78
Слайд 78: Клеточные гистологические элементы
Клетка
Симпласт
Синцитий

Изображение слайда
79
Слайд 79: Типы тканей (Р.А.Кёлликер, Ф.Лейдиг)
Нервная
Эпителиальная
Мышечная
Соединительная (внутренней среды)

Изображение слайда
80
Слайд 80: Типы возбудимых тканей
Нервная
Мышечная
Железистый эпителий

Изображение слайда
81
Слайд 81: При действии раздражителя
В невозбудимой ткани может возникнуть раздражение
В возбудимой ткани может возникнуть раздражение или возбуждение

Изображение слайда
82
Слайд 82: NB! В физиологии возбудимых тканей
Нет понятия «возбудитель»
Есть понятие «раздражитель» (синоним «стимул»)

Изображение слайда
83
Слайд 83: Свойства возбудимых тканей Подробнее – Учебник том I, С.2 7
Вопрос 15

Изображение слайда
84
Последний слайд презентации: Возбудимые ткани. Биоэлектрические явления в возбудимых тканях и методы их: Свойства возбудимых тканей
Возбудимость
Проводимость
Автоматизм
Специфический ответ (сократимость, секреция)

Изображение слайда

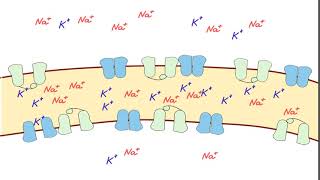
Химический состав живых клеток отличается от внешней среды, причем различия есть не только в сложных молекулах, таких как белки и нуклеиновые кислоты, но и в ионах. Например, во внеклеточной среде преобладают ионы натрия, а в клетке — ионы калия, причем последних на порядок больше. Сама по себе плазматическая мембрана клеток практически непроницаема для ионов, и поэтому для их переноса через мембрану существуют специальные транспортные механизмы — встроенные в мембрану белки. В геноме человека более 800 генов ионных каналов и транспортеров, а общую долю генов, вовлеченных в трансмембранный транспорт, оценивают в 10 % от всех генов, кодирующих белки[1]. В этой серии статей мы рассмотрим механизмы трансмембранного переноса ионов и разнообразие реализуемых ими клеточных функций. Мы также уделим внимание патологиям, вызванным мутациями в генах, кодирующих соответствующие каналы и транспортеры.
Представим себе электрохимическую ячейку — сосуд, разделенный пополам полупроницаемой мембраной, в левой части которого находится 1,0 М раствор KCl, а в правой — 0,1 М KCl. Через мембрану могут проходить катионы K+, но не анионы Cl−. Ионы K+ в результате процесса диффузии будут переходить из левого отсека в правый по градиенту концентрации*, тогда как ионы Cl−, неспособные последовать за катионами, останутся в исходном отсеке. Благодаря такому разделению зарядов на мембране будет накапливаться электрохимический потенциал: избыток анионов с левой стороны мембраны и избыток катионов с правой. Этот потенциал можно измерить, опустив в отсеки электроды, подсоединенные к вольтметру.
Асимметричный поток катионов не будет продолжаться бесконечно: накопленный электрический потенциал (с избытком положительного заряда с левой стороны мембраны) будет противодействовать диффузии ионов калия в левый отсек. Через некоторое время поток ионов K+ из правого отсека в левый сравняется по скорости с потоком из левого отсека в правый, и система достигнет равновесия. Для математического описания подобного равновесия применяют уравнение Нернста (рис. 1).
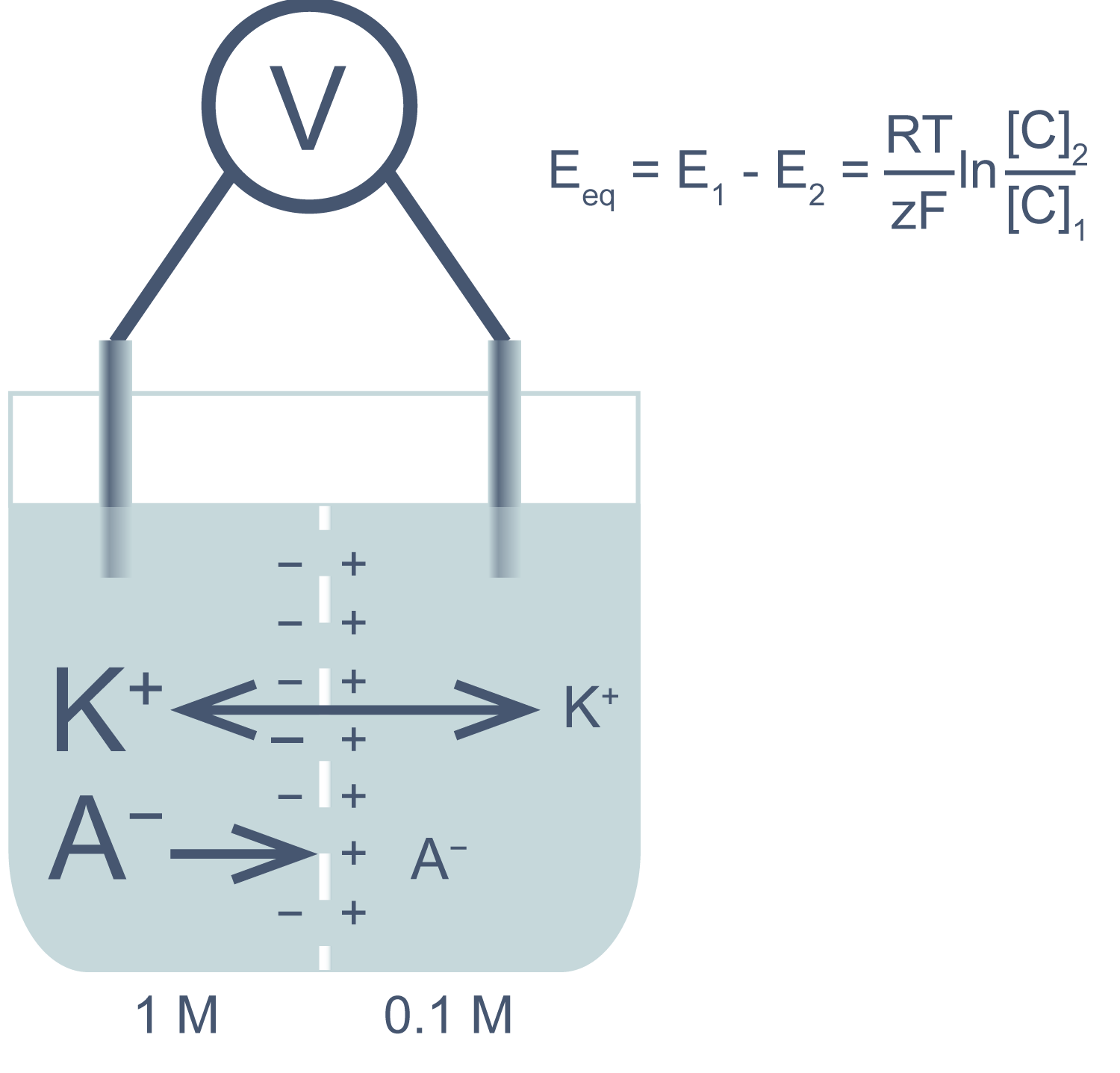
Рисунок 1 | Электрохимическая ячейка. V — вольтметр. Справа приведено уравнение Нернста, где Eeq — равновесный потенциал; E1 – E2 — разность потенциалов по обе стороны мембраны; R = 8,314 Дж/(моль·K) — универсальная газовая постоянная; T — абсолютная температура (в кельвинах); F = 96485,55 Кл·моль–1 — константа Фарадея; z — степень окисления иона (его заряд); [C]1, 2 — равновесные концентрации ионов по обе стороны мембраны.
Если принять, что равновесные концентрации ионов K+ в нашем примере равны начальным, разность потенциала на мембране при 25 °C приблизительно равна –58 мВ.
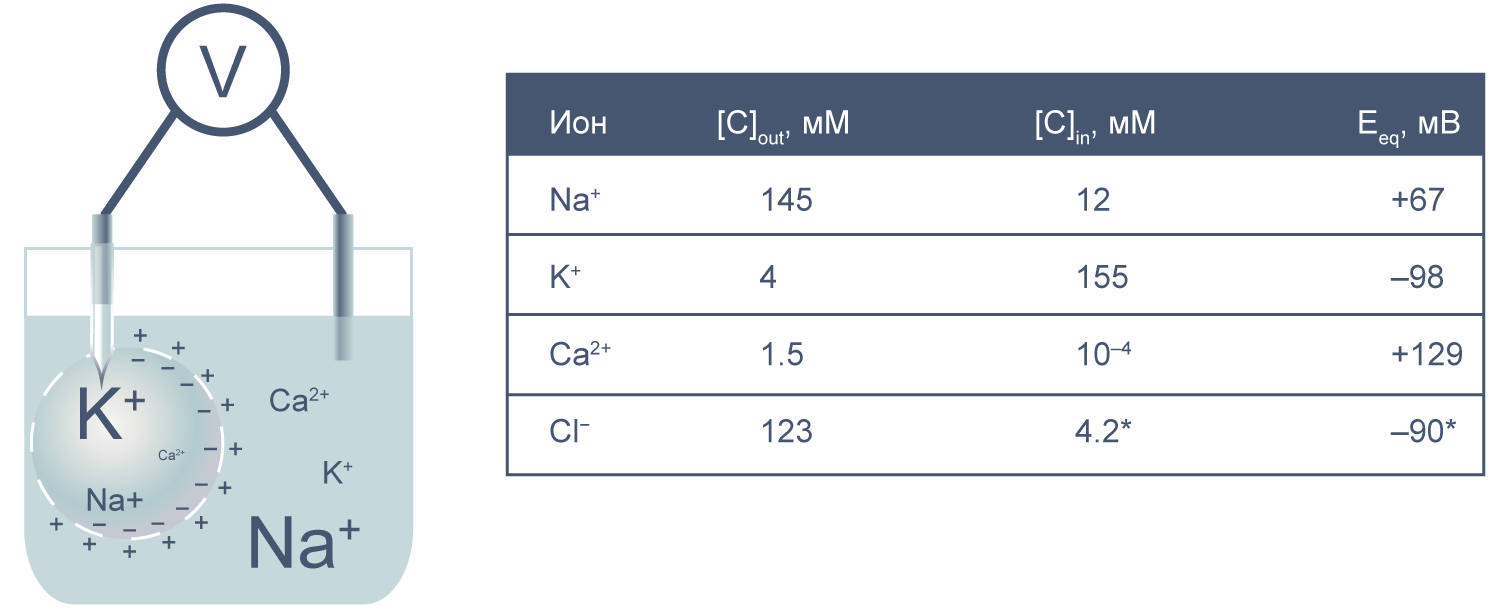
Рисунок 2 | Клетка как электрохимическая ячейка. Справа приведены концентрации основных ионов внутри и вне клетки[2].
Теперь представим, что левая часть нашей электрохимической ячейки — это живая клетка, а правая — внешняя среда. Добавим к этой картине концентрации других физиологически значимых ионов. На мембране клетки также будет накапливаться электрохимический потенциал. Величину электрической составляющей мембранного потенциала измеряют относительно потенциала вне клетки, принимая его за ноль.
В первом приближении можно сказать, что мембрана клетки проницаема для калия и непроницаема для других катионов (Na+, Ca2+) и анионов (в первую очередь для Cl– и отрицательно заряженных участков макромолекул). Ионы калия, выходя из клетки, создают потенциал покоя. Его величина достаточно близка к значению равновесного потенциала для K+, однако строго не равна ему, поскольку в реальности другие катионы и Cl– могут участвовать в формировании потенциала покоя в различных типах клеток. Вычислив равновесные потенциалы для основных ионов, мы получим динамический диапазон величины потенциала на мембране клетки: он не может быть более отрицательным, чем EK, и не может достигать более положительных значений на пике потенциала действия, чем ECa. Причина такого поведения кроется в том, что система стремится к равновесию, и при малейших отклонениях мембранного потенциала в сторону более отрицательных значений, чем EK, K+ будет двигаться по электрохимическому градиенту внутрь клетки, возвращая мембранный потенциал к равновесному потенциалу для калия.
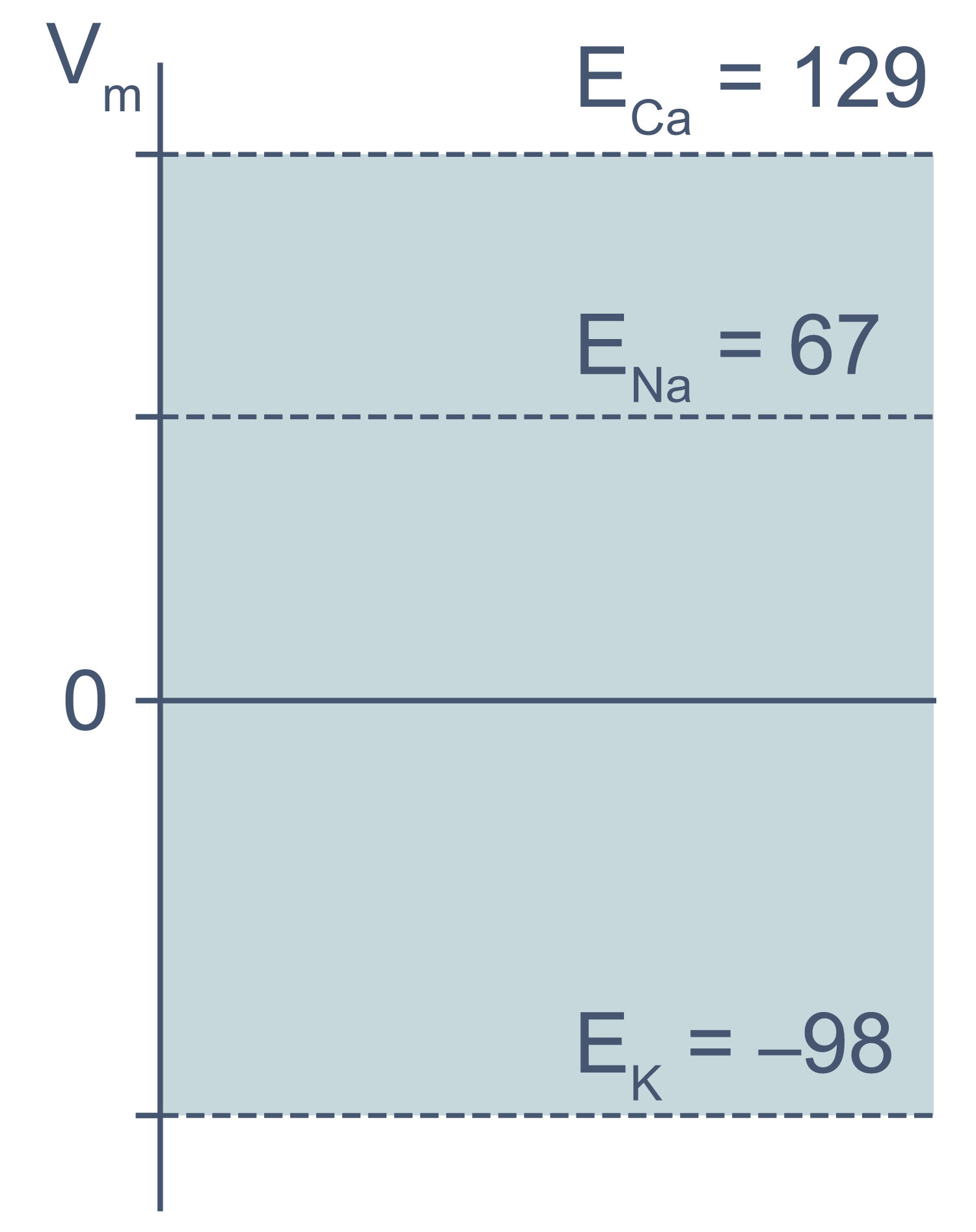
Рисунок 3 | Диапазон возможных значений мембранного потенциала от ЕK до ECa (показан голубым цветом).
Величина потенциала покоя зависит от типа клеток и равна около –30 мВ в невозбудимых клетках и около –80 мВ в возбудимых клетках (нейроны, мышечные и эндокринные клетки). Когда мембранный потенциал более отрицателен, чем потенциал покоя, говорят, что мембрана гиперполяризована, а когда он приближается к нулю или даже принимает положительные значения, говорят о деполяризации мембраны.
В общем случае мембранный потенциал можно вычислить согласно уравнению Гольдмана-Ходжкина-Катца, которое принимает в расчет все основные катионы и анионы:
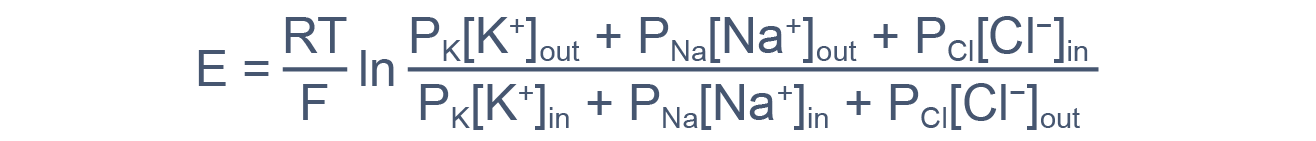
где E — мембранный потенциал; R = 8,314 Дж/(моль·K) — универсальная газовая постоянная; T — абсолютная температура; F = 96485,55 Кл·моль–1 — константа Фарадея; PX — проницаемость мембраны для иона X; [C]in, out — равновесные концентрации ионов внутри и вне клетки. N.B.: для анионов внеклеточная концентрация стоит в знаменателе, а внутриклеточная — в числителе.
Что же обеспечивает проницаемость мембраны для ионов? Заряженные частицы не могут самостоятельно пересекать гидрофобный внутренний слой плазматической мембраны, и поэтому требуются специальные белки, образующие гидрофильную пору, через которую ионы могут двигаться через мембрану. Такие белки называются ионными каналами. Основной вклад в поддержание потенциала покоя вносят калиевые каналы семейств Kir (inward rectifying K+ channels — калиевые каналы внутреннего выпрямления) и K2P (two—pore domain K+ channels — калиевые каналы с двумя поровыми доменами, которые часто называют каналами утечки), а каналы других семейств могут обеспечивать быстрое изменение мембранного потенциала в возбудимых клетках. Каналы могут селективно пропускать определенный тип ионов, например, K+ (как каналы семейства Kir), или более широкий спектр веществ, как, например, коннексины — белки щелевых контактов.
Каналы бывают потенциал-зависимые (потенциал-управляемые), лиганд-зависимые, термо- и механочувствительные — в зависимости от стимула, который управляет открытием и закрытием канала. В роли стимула, таким образом, могут выступать изменения мембранного потенциала, химические агенты, температура, свет, механические и другие стимулы. Один и тот же канал может открываться под действием различных эндо- и экзогенных стимулов. Так, канал TRPV1 активируется повышением температуры более 43 °C[3], кислым pH[4] и разнообразными химическими веществами: капсаицином (алкалоид из перцев рода Capsicum)[3], эндоканнабиноидом анандамидом[5], окситоцином[6] и др.
Физиологическая роль каналов крайне важна. К примеру, мутации в генах, кодирующих белки ионных каналов, лежат в основе патогенеза многих заболеваний человека: некоторых видов эпилепсии[7], муковисцидоза[8], некоторых аритмий[9, 10] и др. Ионные каналы служат мишенями действия многих лекарств, ядов и токсинов.
Однако для формирования потенциала покоя недостаточно одних лишь каналов, ведь нужно создавать и поддерживать концентрационные градиенты на мембране. Основной механизм поддержания градиентов концентрации калия и натрия — это Na/K-АТФаза, фермент, за счет гидролиза одной молекулы АТФ переносящий три Na+ наружу и два K+ внутрь клетки. Она осуществляет электрогенный транспорт: в каждом транспортном цикле при переносе одного дополнительного положительного заряда наружу генерируется некоторая разность потенциалов на мембране. Чтобы оценить этот вклад Na/K-АТФазы в поддержание потенциала покоя, можно заблокировать работу фермента алкалоидом оубаином. Тогда мембрана деполяризуется примерно на 10 мВ[11].
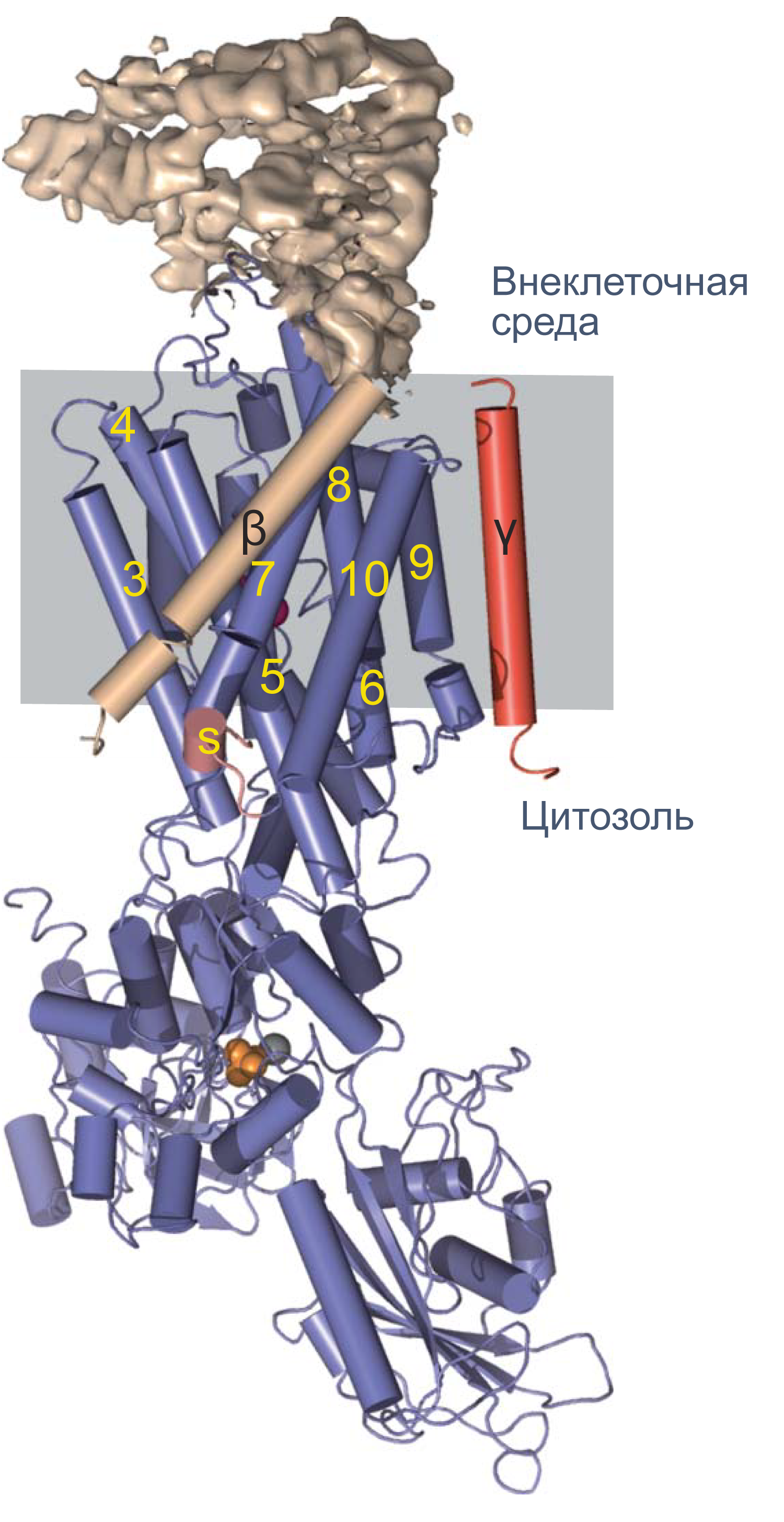
Рисунок 4 | Структура Na/K-АТФазы, полученная методом рентгеновской кристаллографии. Синим цветом показана α-субъединица, красным — γ-субъединица; β-субъединица показана бежевым цветом. Внеклеточная часть β-субъединицы показана в виде электронной плотности. Альфа-спирали изображены цилиндрами, бета-слои — плоскими стрелками[12].
Активный и пассивный транспорт веществ через мембрану
Процессы транспорта веществ через мембрану можно классифицировать по источнику энергии для транспорта. Пассивный транспорт — это движение вещества через канал или транспортер по градиенту концентрации*, то есть за счет энергии электрохимического градиента. Таким способом через калиевые каналы пассивно движутся ионы калия, или осуществляется перенос глюкозы через транспортер GLUT4 (такой тип транспорта еще называют облегченной диффузией, а транспортеры, переносящие только один субстрат — унипортерами). Кроме пассивного транспорта, существует активный транспорт, при котором субстраты переносятся против градиента концентрации с затратой энергии, запасенной клеткой в виде АТФ (например, Na/K-АТФаза).
Некоторые транспортеры сопрягают перенос ионов или молекул против градиента концентрации с движением ионов по градиенту концентрации. Симпортеры переносят различные частицы в одном направлении (например, KCC2 — K—Cl cotransporter 2 — K/Cl котранспортер 2), а антипортеры, или обменники, — в противоположных (например, NHE-1 — Na/H exchanger 1 — Na/H обменник, участвующий в поддержании клеточного pH). Такой транспорт называется вторично-активным.
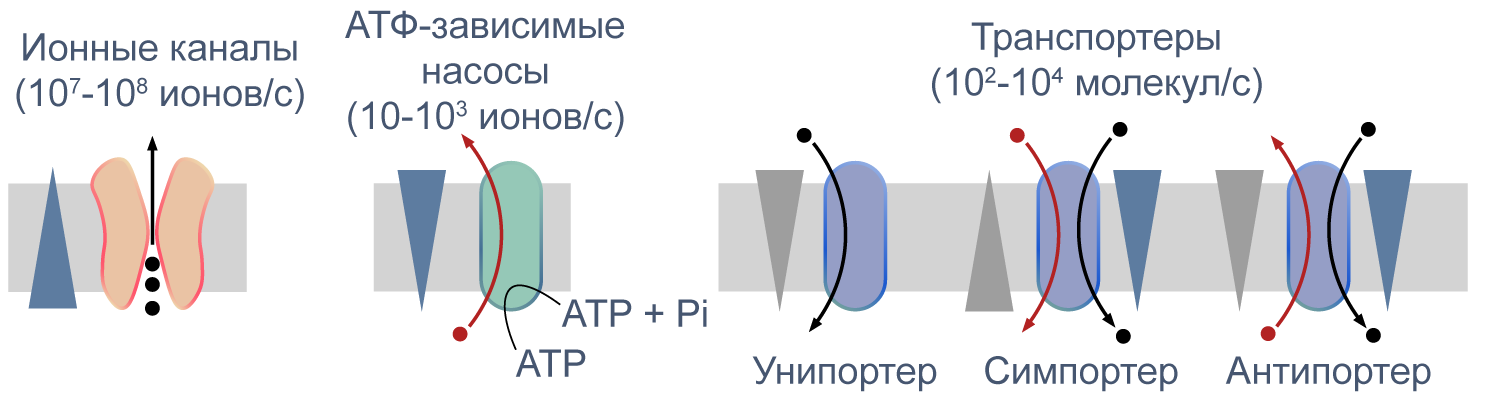
Рисунок 5 | Пассивный и активный транспорт веществ через мембрану. Треугольниками показаны концентрационные градиенты. Черными стрелками показано движение по градиенту концентрации, красными — против градиента концентрации. По [13], с изменениями.
В старой литературе можно встретить концепцию «белков-переносчиков»: до открытия молекулярной идентичности многих каналов и транспортеров (т. е. какая молекула опосредует данный ионный ток и какой ген ее кодирует) существовало представление о переносчиках как о челноках, связывающих субстраты с одной стороны мембраны, диффундирующих через мембрану и высвобождающих субстраты с другой стороны. Однако, когда стали известны аминокислотные последовательности транспортных белков, стало понятно, что все они часто содержат многочисленные трансмембранные домены и образуют сквозной путь через мембрану.
Различия между каналами, транспортерами и насосами заключаются в механизме их работы и регуляции. Каналы представляют собой более или менее селективную пору, через которую ионы могут свободно диффундировать, не вызывая конформационных изменений в белке канала. При токе через отдельный канал в 1 пА по нему проходят 6×106 одновалентных ионов в секунду. Транспортер, связываясь с субстратом, изменяет свою конформацию для переноса субстрата. При ко-транспорте повышается аффинность транспортера в новой конформации к второму субстрату, и субстраты переносятся через мембрану сопряженно. Насосы, также называемые АТФазами, (ауто)фосфорилируются АТФ, и это фосфорилирование значительно изменяет их конформацию и приводит к транслокации субстратов через мембрану. Электрическая проводимость насосов очень мала: Na/K-АТФаза переносит всего около 300 Na+ и 200 K+ в секунду.
Граница между молекулами каналов и транспортеров не всегда строга. Например, семейство хлоридных каналов и транспортеров ClC (англ. Chloride Channel) включает в себя гомологичные друг другу каналы (ClC-1, 2, Ka и Kb) и Cl/H-обменники (ClC-3–7), которые можно превратить в каналы, внеся мутацию в единственный остаток глутамата (т. н. gating glutamate — воротный глутамат) [14]. Na/K-АТФазу также можно превратить в простую пору, например, с помощью палитоксина[15]. Хлоридный канал CFTR относится к группе ABC-транспортеров, однако он использует энергию АТФ не для транспорта ионов, а для регуляции открытия и закрытия канала[16]. Кроме того, существуют данные о том, что везикулярный транспортер глутамата VGLUT1 кроме обмена глутамата на протоны также опосредует не сопряженный с обменом ток Cl− [17].
Клеточная мембрана как электрическая цепь
Липидный бислой мембраны можно представить как резистор (сопротивление) и конденсатор (емкость), соединенные параллельно. Величина сопротивления зависит от плотности каналов в мембране и их функционального состояния. Электроемкость возникает из-за разделения зарядов по обе стороны мембраны тонким слоем диэлектрика (гидрофобными хвостами фосфолипидов).
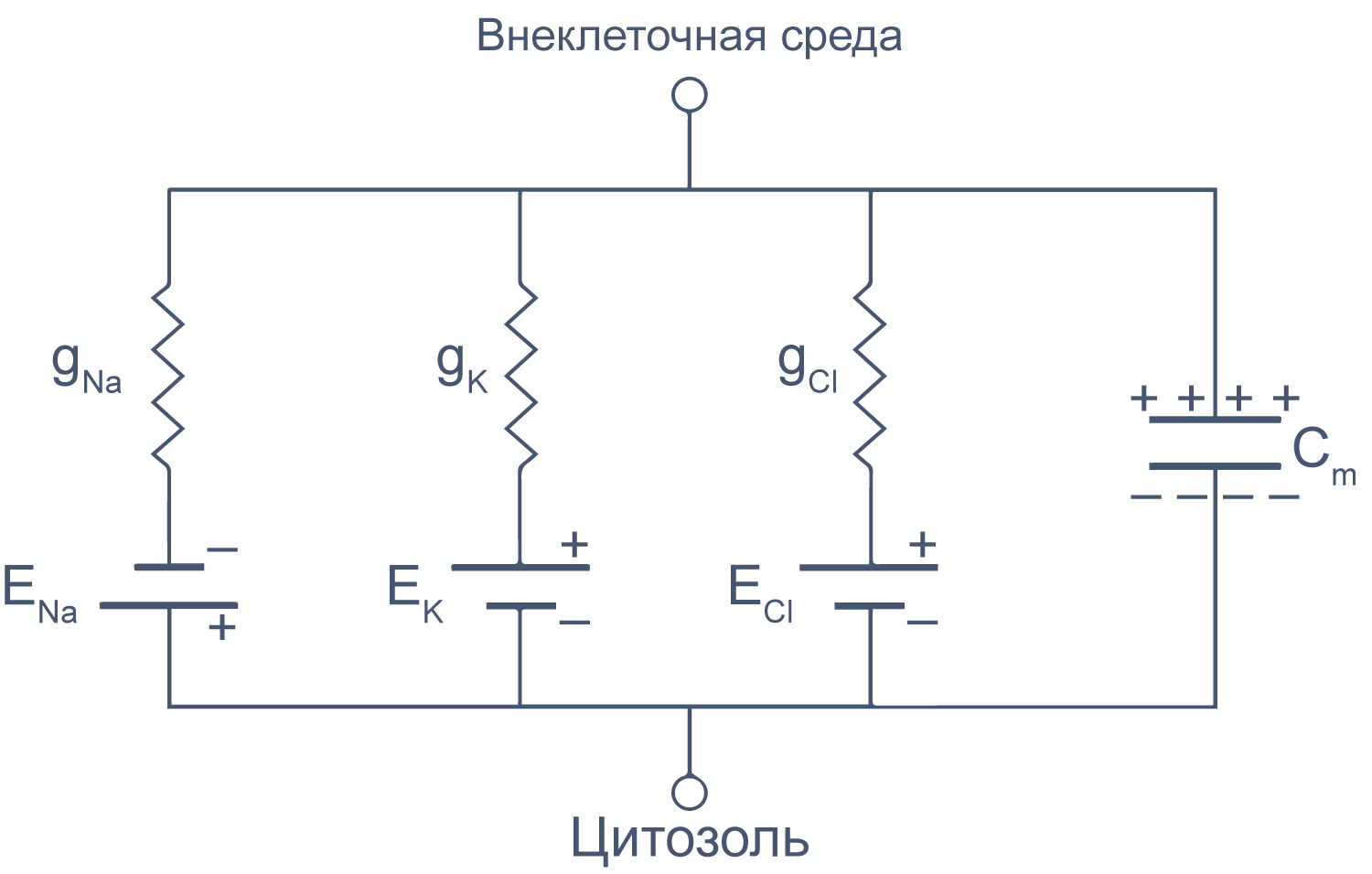
Рисунок 6 | Плазматическая мембрана, представленная в виде RC схемы. gNa, gK, gCl — проводимость** мембраны для ионов Na+, K+ и Cl–, соответственно; ENa, EK, ECl — электродвижущая сила, или равновесные потенциалы для соответствующих ионов, Сm — электроемкость мембраны.
Можно считать, что проводимость (величина, обратная сопротивлению, измеряемая в сименсах) характеризует проницаемые для ионов компоненты структуры мембраны, а емкость — не проницаемые для ионов структуры. Удельная емкость мембраны составляет приблизительно 1 мкФ/см2 или 0,01 пФ/мкм2*** и мало зависит от типа клеток[2]. Это позволяет оценить размер клетки по ее электрическим характеристикам. Высокая электроемкость клеточных мембран задерживает изменения потенциала в ответ на ток. Эту задержку можно выразить через постоянную времени τ, которая равна произведению емкости и параллельно соединенного с ней сопротивления (RC). Если конденсатор зарядить на некоторую величину, а затем дать ему разрядиться через резистор, то потенциал будет убывать экспоненциально согласно формуле V = V0e–t/RC, то есть за каждые τ = RC секунд потенциал будет падать в 1/e раз (на 37 % от исходной величины). Этот расчет применим и к клеточной мембране: падение напряжения при разрядке мембраны будет подчиняться экспоненциальному закону. То есть, если приложить гипер- или деполяризующий стимул, то мембранный потенциал через время τ достигнет 63 % конечного сдвига.
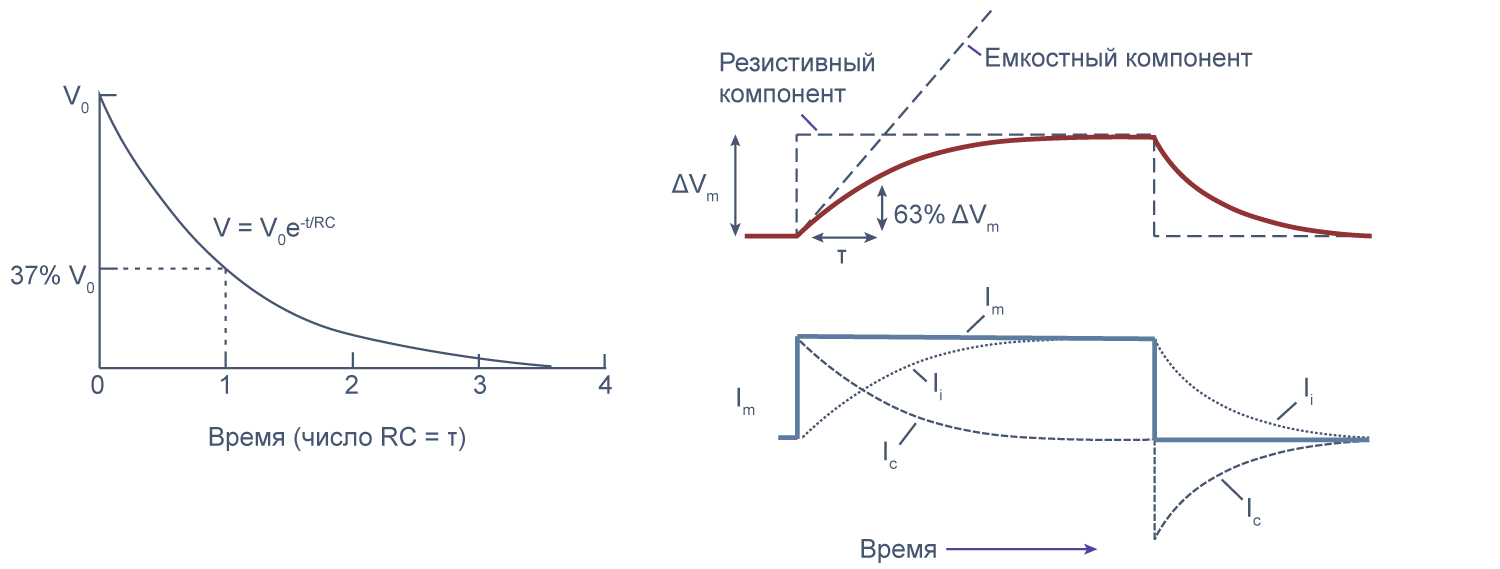
Рисунок 7 | Падение напряжения при разрядке мембраны. Изменение мембранного потенциала при деполяризации мембраны на ΔVm [22], Ic — емкостный ток, Ii — ионный ток, Im — суммарный ток через мембрану. Пояснения в тексте.
Потенциал действия
Возбудимые клетки могут быстро изменять потенциал на мембране и этим запускать внутриклеточные процессы, например, мышечное сокращение или экзоцитоз везикул. Кратковременный локальный скачок мембранного потенциала от приблизительно –80 мВ (значения потенциала покоя) до 0…+20 мВ за счет изменения проницаемости мембраны для ионов называется потенциалом действия.
Для начала рассмотрим потенциал действия в аксонах нейрона на примере гигантского аксона кальмара. Выбор столь экзотического объекта обусловлен историческими причинами. Современная микроэлектродная техника — использование тонких стеклянных микропипеток, заполненных раствором электролита, — была предложена лишь в 1949 году Дж. Лингом и Р. Джерардом[18]. До этого поместить электрод, представлявший собой тонкую проволоку, внутрь клетки, не разрушив ее, можно было только в случае ее крупных размеров. Гигантский аксон кальмара стал идеальным модельным объектом для изучения потенциала действия: его диаметр может достигать 0,5–1 мм. Этот отросток служит для быстрой передачи нервных импульсов у беспозвоночных, у которых отсутствует миелинизация. Внутренним содержимым аксона легко манипулировать, заменяя ионный состав внутриклеточного раствора, а внутрь такого аксона можно поместить электрод для регистрации мембранного потенциала.
Так, в 1939 году А. Ходжкин и Э. Хаксли опубликовали в журнале Nature короткую заметку «Потенциалы действия, зарегистрированные внутри нервного волокна»[19], в которой они описали первый эксперимент по внутриклеточному измерению мембранного потенциала на гигантском аксоне кальмара. Посмотрим на потенциалы действия, которые зарегистрировали исследователи.
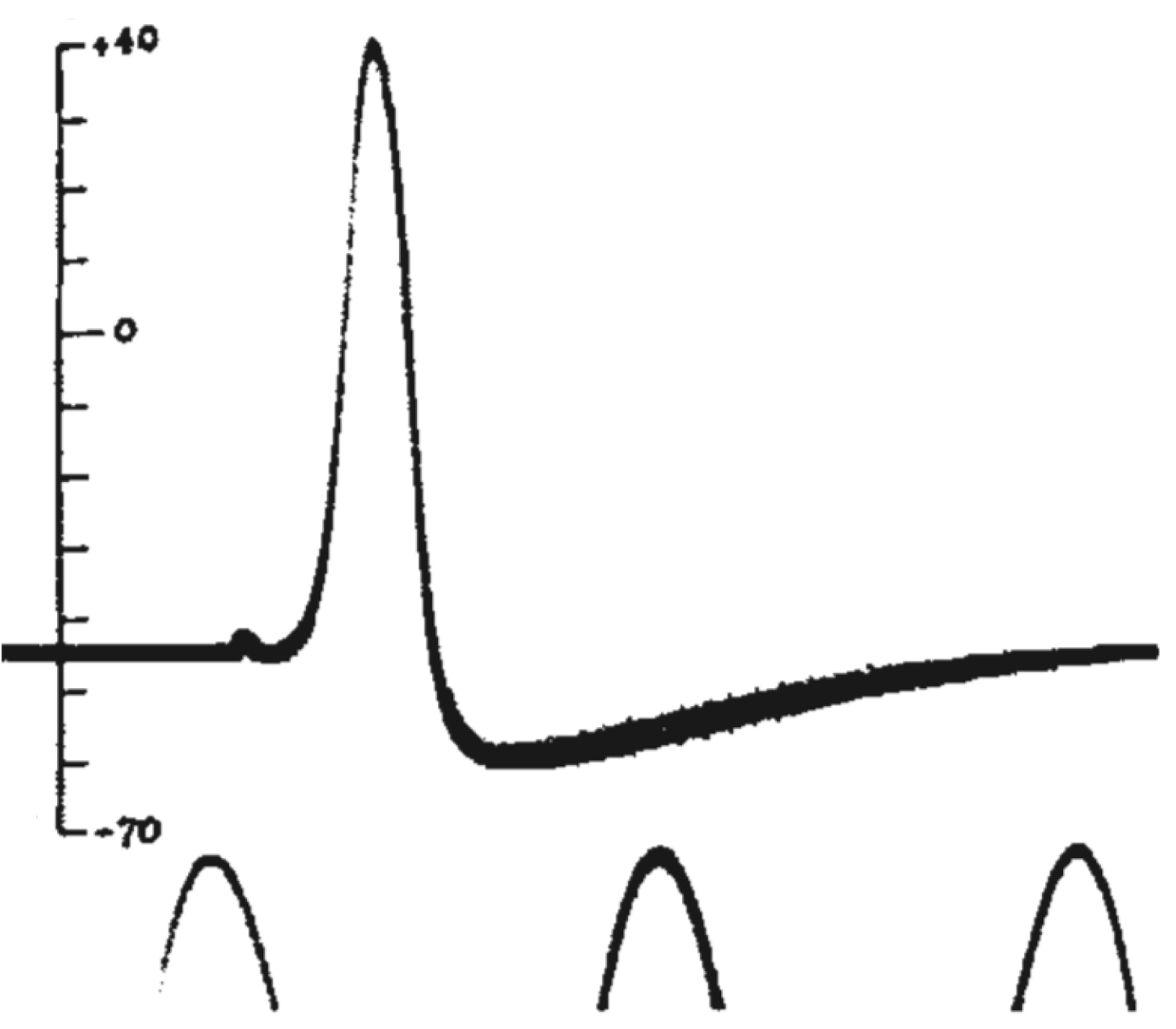
Рисунок 8 | Потенциал действия из статьи А. Ходжкина и Э. Хаксли 1939 года[19]. Потенциал действия был зарегистрирован как разность потенциала между внутренней средой аксона и внешней средой. Пики внизу рисунка — это фрагменты синусоидального сигнала с частотой 500 Гц. Вертикальной чертой отмечен потенциал внутреннего электрода в милливольтах, потенциал морской воды снаружи аксона был принят за ноль.
Мы видим резкую деполяризацию мембраны до положительных значений, а затем более плавное возвращение потенциала к отрицательным значениям, зачастую более отрицательным, чем величина потенциала покоя. Из этих наблюдений следовало два важных вывода: 1) потенциал действия генерируется мембраной клетки, что не было очевидно в 1930-е годы; 2) поскольку измеренная амплитуда потенциала действия была больше величины мембранного потенциала, генерация потенциала действия — это активный процесс, который нельзя объяснить временным «пробоем» в мембране (каким бы наивным такое представление нам не казалось сегодня, в 1930-е так думали многие физиологи). В том же году К. С. Коул и Х. Дж. Кертис[20] зарегистрировали кратковременное изменение проводимости мембраны с 1 мСм/см2 до 40 мСм/см2 при генерации потенциала действия. Эти данные также свидетельствовали о тонкой регуляции этого процесса.
После Второй мировой войны Ходжкин и Хаксли вернулись к своим исследованиям. В 1945 году они опубликовали статью с несколькими дополнительными экспериментами, подтверждающими сформулированные ранее выводы. Но и тогда ионные механизмы потенциала действия оставались неясными.
В 1952 году вышла серия статей, посвященных разгадке механизма генерации потенциала действия. Ведущую роль в этом открытии сыграл новый метод фиксации потенциала с помощью двух электродов (в англоязычной литературе TEVC — two-electrode voltage clamp), разработанный в конце 1940-х годов К. С. Коулом и Дж. Мармонтом[21]. Этот метод позволяет измерять не только потенциал, но и токи при заданном значении мембранного потенциала. Как же осуществляется фиксация потенциала с помощью двух электродов на заданном уровне (Vcmd — «командный потенциал»)? В клетку помещают два электрода, один из которых измеряет потенциал (относительно внеклеточного электрода сравнения) и передает его значение на специальный усилитель, который сравнивает измеренный потенциал со значением командного потенциала. Это устройство вычисляет ток, необходимый для компенсации этой разницы, и подает через второй внутриклеточный электрод ток такой величины, чтобы потенциал на мембране клетки стал равен Vcmd (Vm = Vcmd). Из амплитуды тока, необходимого для компенсации сдвига потенциала до Vcmd, можно сделать вывод о токе через мембрану при данном значении мембранного потенциала. Ток при данном значении потенциала равен току, подаваемому на второй электрод, взятому с обратным знаком.
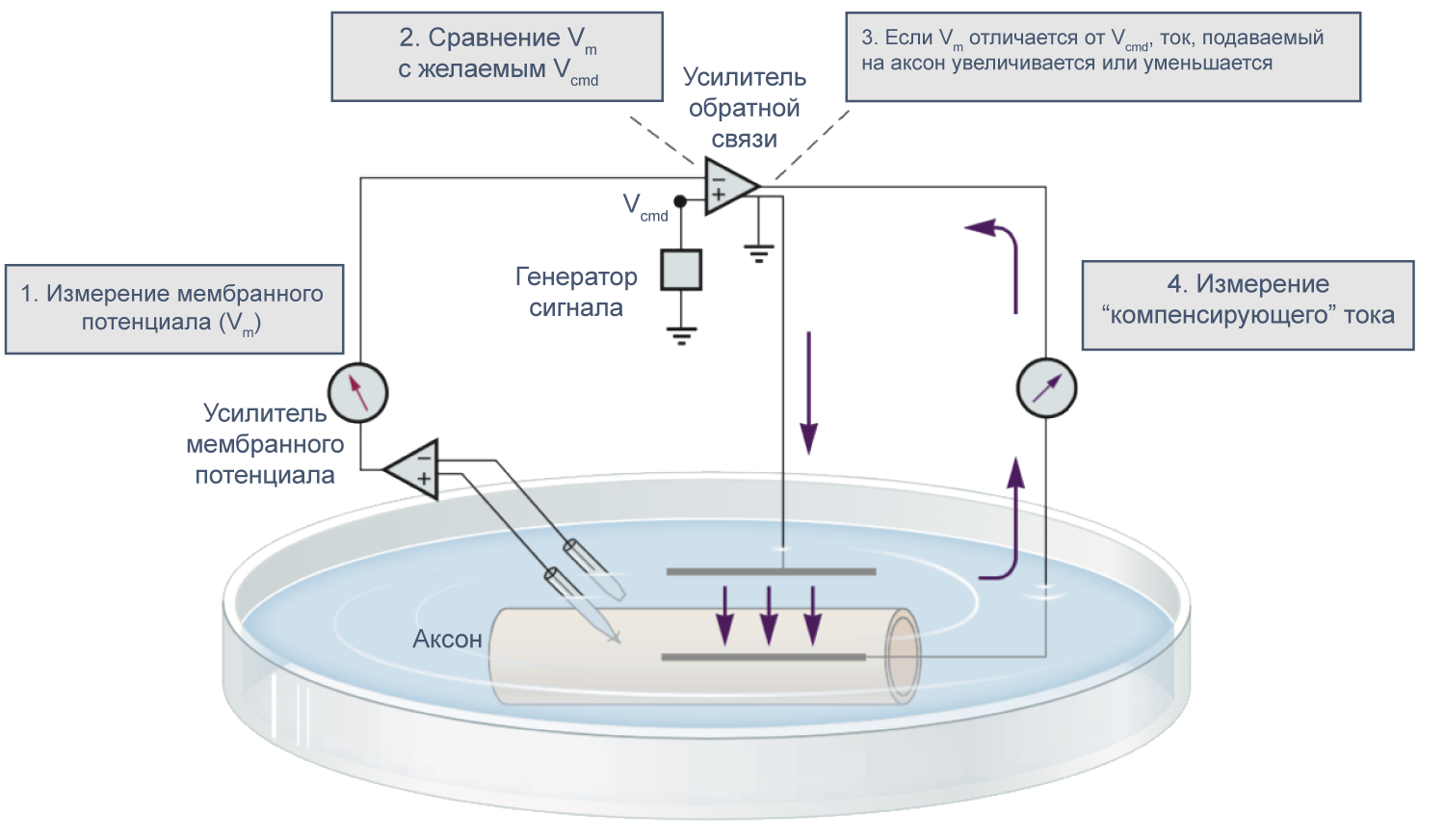
Рисунок 9 | Схема фиксации потенциала с помощью двух электродов[21].
В 1970–80-х годах Эрвин Неер и Берт Сакман (Erwin Neher, Bert Sakmann) предложили метод локальной фиксации потенциала (англ. patch clamp), позволяющий работать с мелкими клетками и токами малой амплитуды и даже регистрировать активность отдельных каналов. Тем не менее, метод фиксации потенциала двумя электродами в микроэлектродной конфигурации используется и сегодня при работе с такими крупными клетками, как ооциты лягушки Xenopus laevis.
Временной ход потенциала действия
Потенциал действия в аксоне можно разделить на 1) фазу быстрой деполяризации до 2) положительных значений (овершута, от англ. overshoot), 3) фазу реполяризации, в которой потенциал возвращается к потенциалу покоя или даже до несколько более отрицательных значений — 4) следовая гиперполяризация.
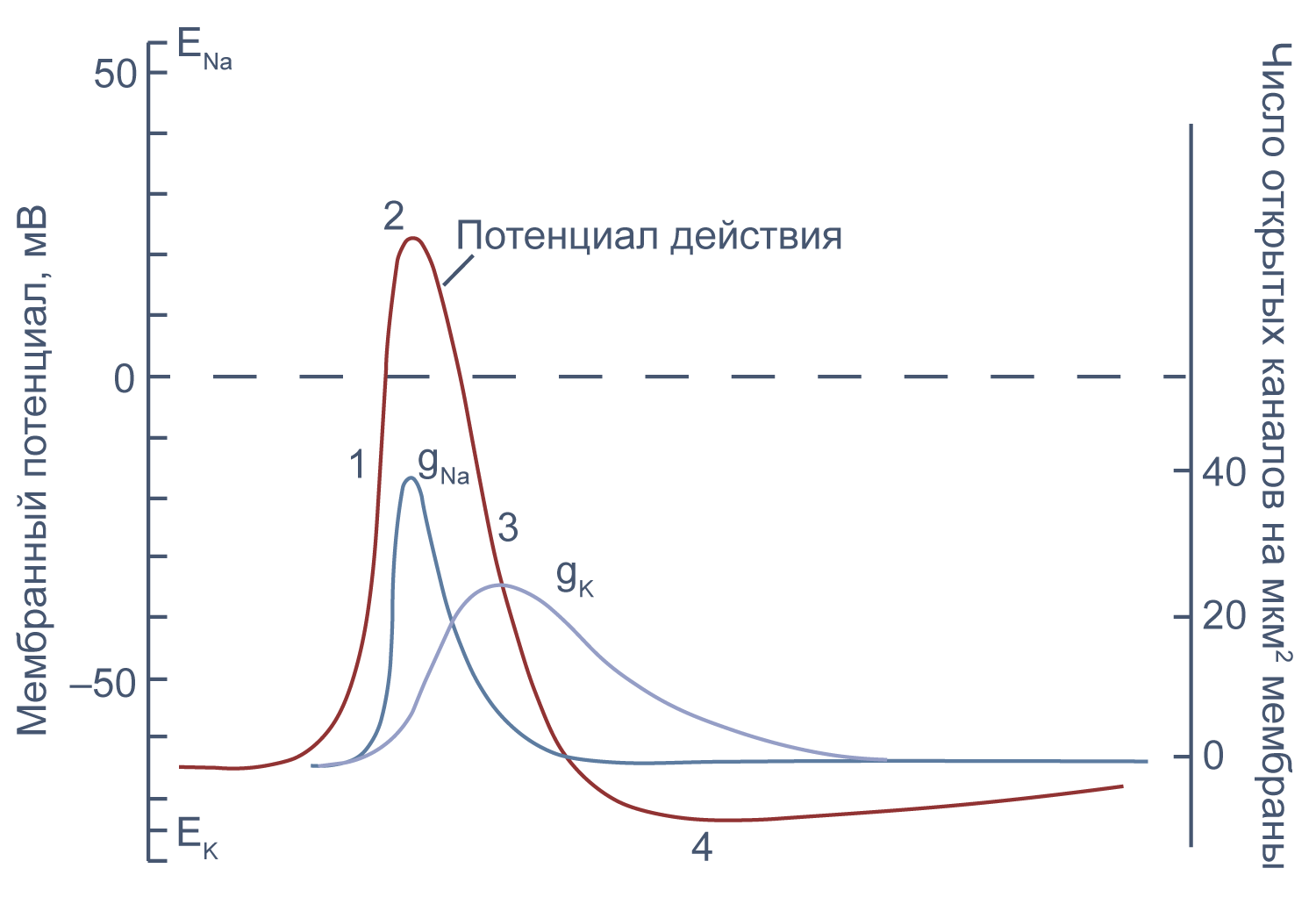
Рисунок 10 | Потенциал действия и изменения проводимости мембраны для Na+ и K+ в гигантском аксоне кальмара. Из [22], с изменениями.
Изменяя ионный состав внутри- и внеклеточного раствора, можно изолировать ионные токи, которые опосредуют изменение мембранного потенциала при потенциале действия. Так, помещая аксон в раствор, в котором натрий заменен на холин, можно изолировать натриевый и калиевый компонент потенциала действия, то есть отдельно измерить калиевый ток[20]. Этого же можно достичь применением блокаторов потенциал-зависимых натриевых и калиевых каналов — тетродотоксина и тетраэтиламмония (TEA).
Чтобы понять взаимосвязь ионных токов и вызванных ими изменений потенциала, рассмотрим всю цепь событий при генерации потенциала действия. Сначала мембрана деполяризуется под действием внешнего стимула: поступления в клетку катионов через лиганд-управляемые каналы, закрытия калиевых каналов или электрической стимуляции в эксперименте. Если деполяризация достигает порогового значения для потенциалзависимых натриевых каналов (Nav), они открываются, натрий по градиенту своей концентрации входит в клетку, и мембрана деполяризуется еще сильнее. Дальнейшая деполяризация влечет за собой лавинообразное открытие все новых натриевых каналов, ток через которые приводит к еще большей деполяризации. Однако эта петля положительной обратной связи не работает бесконечно: открывшись на некоторое время, натриевые каналы инактивируются и не могут открыться вновь, пока мембранный потенциал не вернется к отрицательным значениям.
Механизм инактивации был предложен еще в математической модели Ходжкина и Хаксли[23] на основании кинетических характеристик тока. Они предположили, что в натриевом канале есть три активационные частицы m и одна инактивационная частица h. Когда стала известна аминокислотная последовательность канала, выяснилось, что канал на самом деле имеет четыре гомологичных активационных домена и один инактивационный, однако один из активационных доменов срабатывает значительно медленнее остальных трех, и его влияние на кинетические характеристики тока маскируется происходящей в то же время инактивацией канала (например, [24]).
В то же время деполяризация мембраны приводит к активации потенциал-зависимых калиевых каналов (Kv), которые открываются медленнее, чем натриевые, калий выходит из клетки, и потенциал возвращается к потенциалу покоя и может даже временно стать более отрицательным: пока натриевые каналы инактивированы, потенциал становится ближе к калиевому равновесному потенциалу, и это явление называется следовой гиперполяризацией.
Генерация потенциала действия происходит по принципу «все или ничего». Если деполяризующий стимул не достиг порогового значения, потенциал действия не генерируется. Если же порог был достигнут, положительная обратная связь обеспечивает открытие всех доступных натриевых каналов, и потенциал действия достигает своей максимальной амплитуды.
Если новый стимул приходит во время или сразу после генерации потенциала действия, второй потенциал действия не возникает или обладает меньшей амплитудой, чем первый. Это явление называется рефрактерностью. Стимул, возникший в период абсолютной рефрактерности, не вызывает генерации потенциала действия, а пришедшийся на период относительной рефрактерности вызывает потенциал действия уменьшенной амплитуды, так как часть натриевых каналов все еще инактивирована.
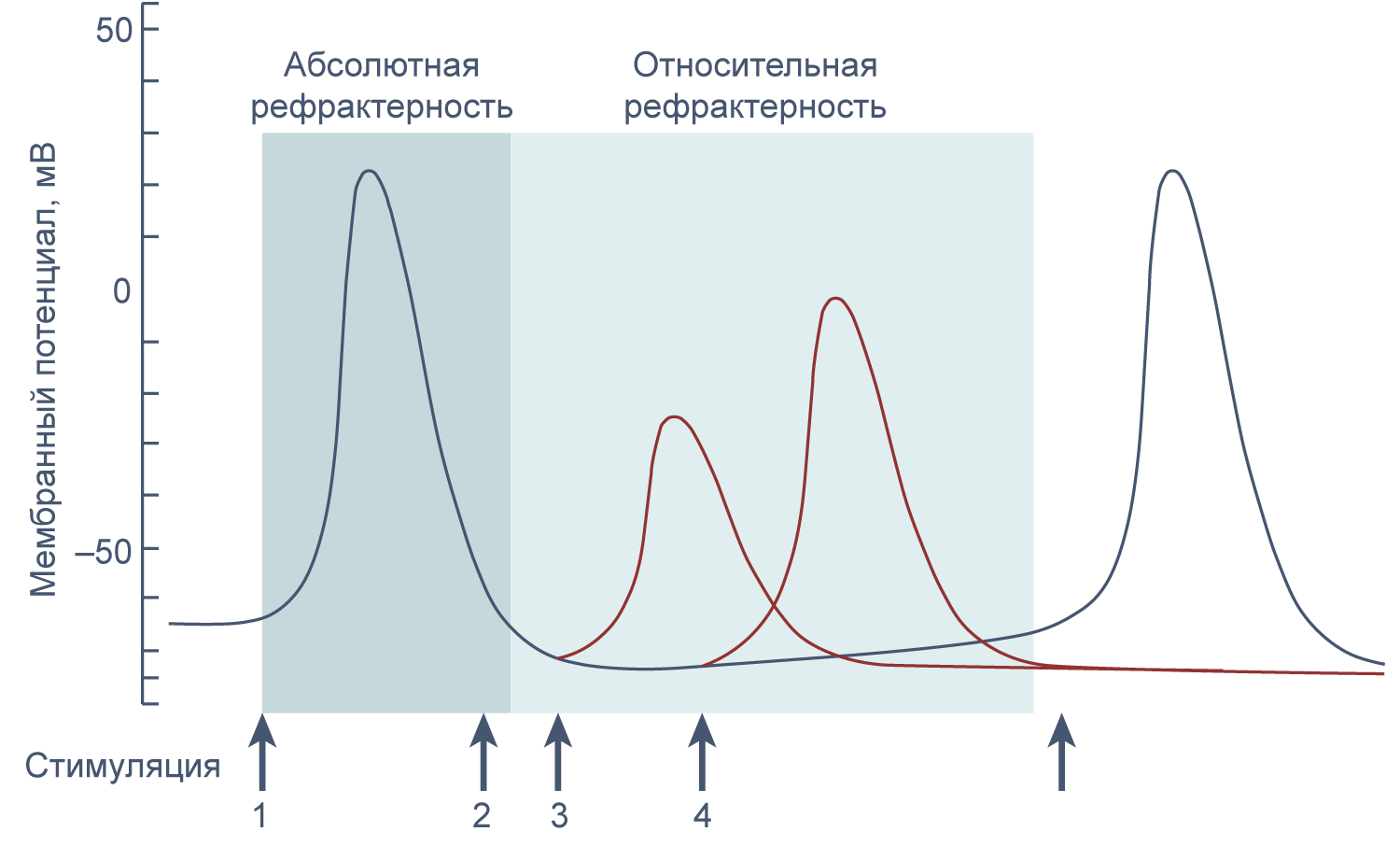
Рисунок 11 | Абсолютная и относительная рефрактерность. Стимуляция в период абсолютной рефрактерности (2) не ведет к генерации потенциала действия, а стимуляция в период относительной рефрактерности (3 и 4) приводит к генерации потенциала действия сниженной амплитуды.
Стоит отметить, что внутриклеточная концентрация физиологически значимых ионов при генерации потенциала действия не меняется, и ионные токи задействуют пренебрежимо малую долю от общего числа Na+ и K+. Это можно проиллюстрировать следующим примером. Рассчитаем число ионов, которое должно пересечь мембрану для деполяризации на 100 мВ. Заряд на мембране равен произведению емкости мембраны и потенциала: Q = CmVm. Удельная емкость мембраны близка к 1 мкФ/см2, а сдвиг потенциала в нашем случае равен 0,1 В. Тогда количество разделенных зарядов равно Q = 10−6 Ф/см2 × 0,1 В = 10−7 Кл/см2. Величина заряда одного иона Na+ или K+ (элементарного заряда) равна 1,6 × 10−19 Кл, тогда количество переносимых через мембрану ионов равно 10−7 Кл/см2 / 1,6 × 10−19 Кл = 6,25 × 1011 ионов/см2, или 6250 ионов/мкм2. Для клетки диаметром 10 мкм площадь поверхности мембраны будет приблизительно равна 4πr2 = 314 мкм2 (в этом расчете для простоты мы считаем клетку гладкой сферой), а объем — 4πr3/3 = 524 мкм3. При внутриклеточной концентрации Na+ 10 мМ, а K+ 150 мМ содержание этих ионов в цитозоле будет равно 3,2 × 109 и 4,7 × 1010 соответственно. В течение одного потенциала действия приблизительно 314 мкм2 × 6,250 ионов/мкм2 ≈ 2 000 000 ионов Na+ входит в клетку в фазе деполяризации и примерно столько же ионов K+ выходит из клетки в фазе реполяризации, что составляет всего 0,06 % от общего числа ионов натрия в клетке. Токи такой величины обычно не изменяют макроскопические концентрации ионов в клетке, поскольку работа Na/K-АТФазы компенсирует эти незначительные изменения. Однако при определенных условиях концентрации Na+ и K+ все же могут измениться, например, при продолжительной стимуляции аксонов с малым диаметром[25].
Распространение возбуждения по аксону
Электрические свойства мембраны помогают понять изменения мембранного потенциала не только во времени, но и в пространстве. Пассивное, или электротоническое распространение возбуждения по мембране происходит без изменения проводимости потенциал-зависимых каналов. Для распространения потенциала действия важен как активный (изменение проводимости натриевых и калиевых каналов), так и пассивный механизмы, поскольку деполяризация, вызывающая открытие новых потенциал-зависимых натриевых каналов, должна достигнуть нового невозбужденного участка аксона, и происходит это благодаря электротоническому распространению возбуждения.
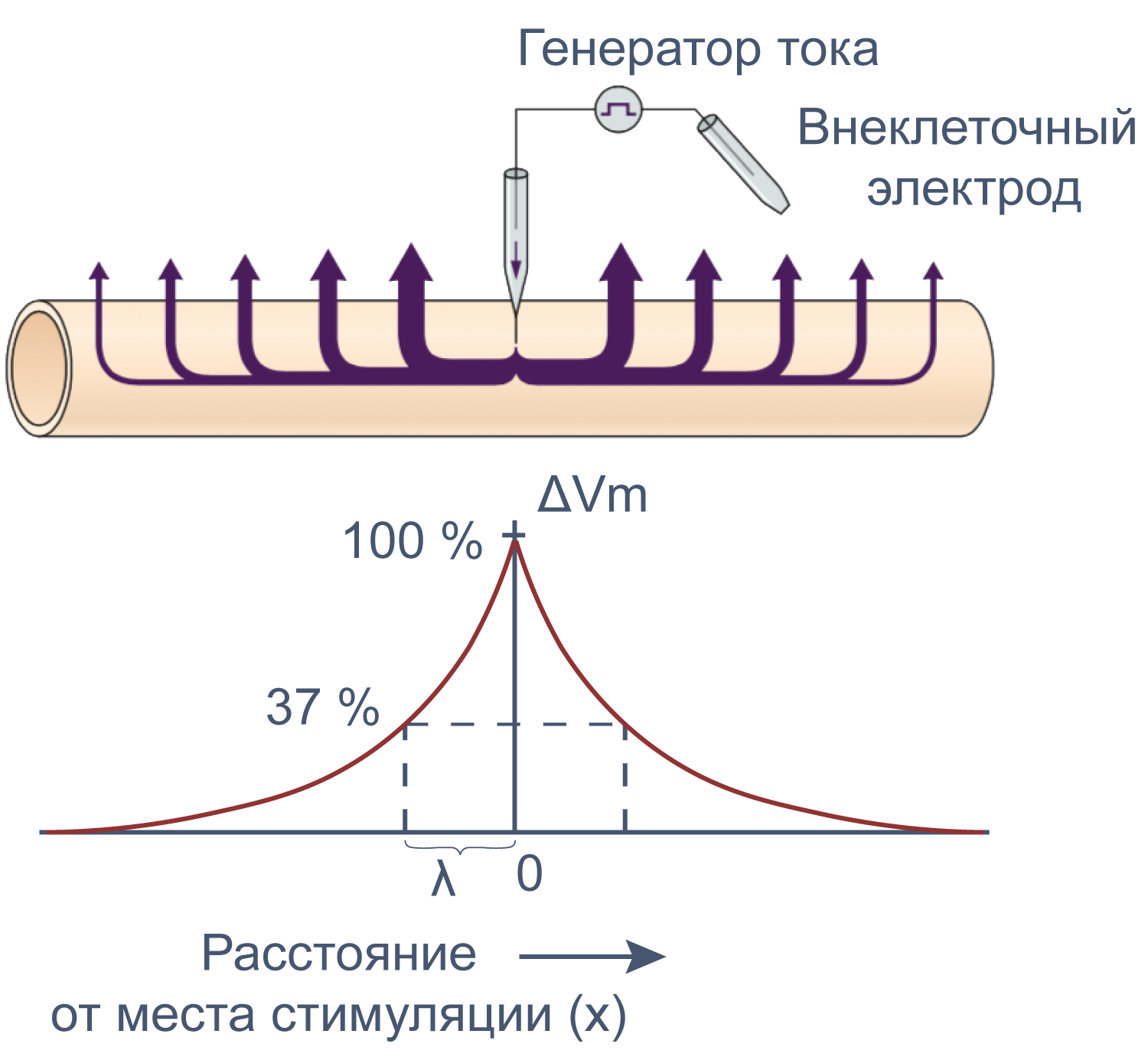
Рисунок 12 | Электротоническое распространение возбуждения. Пояснения в тексте [22].
Сдвиг потенциала в точке, отстоящий на x от места стимуляции, можно вычислить как Ex = E0e–x/λ, где E0 — это сдвиг потенциала в точке стимуляции, а λ — постоянная длины.
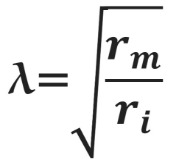
Постоянная длины возрастает с увеличением сопротивления мембраны (rm) и уменьшается с возрастанием сопротивления аксоплазмы (ri), которое в свою очередь зависит от концентрации подвижных зарядов в объеме аксона. Знание этих закономерностей позволяет понять зависимость скорости распространения возбуждения от радиуса (R) аксона. Сопротивление мембраны rm пропорционально 1/2πR, сопротивление аксоплазмы ri — 1/πR2, а емкость мембраны Cm пропорциональна R. По мере увеличения радиуса аксона и rm, и ri уменьшаются, но ri уменьшается сильнее. Постоянная длины увеличивается, следовательно, сдвиг потенциала может распространяться по более крупному аксону дальше. Кроме того, увеличение радиуса аксона ведет к увеличению емкости мембраны, однако этот эффект нивелируется тем, что емкость с увеличением радиуса растет линейно, а уменьшение ri пропорционально квадрату радиуса. Таким образом, проводимость аксоплазмы увеличивается быстрее (при снижении ri), чем растет емкость мембраны, и это позволяет току быстрее достигать невозбужденных участков мембраны более крупного аксона.
Зависимость скорости распространения потенциала действия по аксону от его диаметра объясняет необходимость в гигантских аксонах у кальмара. Однако у позвоночных гигантских аксонов нет, и достаточная скорость проведения нервных импульсов достигается с помощью миелинизации аксонов. Миелиновые оболочки образованы специальными глиальными клетками: олигодендроцитами в центральной нервной системе и клетками Шванна в периферической. Эти клетки оборачивают аксон, образуя вокруг него плотный чехол из десятков слоев плазматической мембраны, который работает как изолятор. Под миелиновой оболочкой очень мало или почти нет натриевых каналов. Они оказываются расположены на небольших участках мембраны аксона между двумя соседними шванновскими клетками или олигодендроцитами — в перехватах Ранвье. Миелиновая оболочка увеличивает скорость проведения потенциала действия благодаря повышению сопротивления мембраны rm. При этом снижаются потери тока через мембрану, и деполяризация может пассивно распространяться на бо́льшие расстояния. Кроме того, из-за снижения емкости мембраны Cm, меньшая доля тока тратится на перезарядку мембраны. Миелинизация делает передачу потенциала действия не только быстрее, но и эффективнее, поскольку натриевые каналы располагаются только в перехватах Ранвье, благодаря чему меньшее количество ионов натрия входит в клетку и меньше энергии требуется на работу Na/K АТФ-азы для поддержания концентрационного градиента.
Потенциалы действия в других возбудимых клетках
Минималистичный натриево-калиевый потенциал действия характерен только для аксонов нейронов. В других частях нейрона и в иных типах возбудимых клеток в генерации потенциала действия принимают участие разнообразные ионные каналы, в том числе калиевые каналы других семейств и кальциевые каналы плазматической мембраны и эндоплазматического ретикулума (ЭПР является внутриклеточным депо кальция; общая концентрация Ca2+ в нем достигает миллимолярных значений, а концентрация свободного кальция находится в микромолярном диапазоне[26]; эти значения на несколько порядков выше, чем 100 нМ свободного Ca2+ в цитозоле в состоянии покоя). Различия в экспрессии генов ионных каналов в различных типах возбудимых клеток порождают разнообразие потенциалов действия, различающихся ионными механизмами, длительностью (от 1,5 мс в аксонах до 500 мс в кардиомиоцитах желудочков), необходимостью внешнего стимула для генерации или наличием собственного ритма. Многие ионные каналы, не участвуя напрямую в генерации потенциала действия, влияют на возбудимость клеток и таким образом вносят вклад в разнообразие электрической активности клеток.
* Принятое в физиологической литературе употребление фраз «по» или «против градиента концентрации» расходится с физическим понятием градиента. В математике и физике градиент направлен в сторону наибольшей скорости возрастания функции или величины; так, если вещество движется в направлении этого вектора, то физиологи говорят о движении «против градиента концентрации», а если в противоположном направлении — то «по градиенту концентрации». Такое словоупотребление прочно закрепилось в литературе, но пусть оно не сбивает вас с толку, когда вы размышляете, откуда взялся минус перед градиентом концентрации в уравнении диффузионного потока (уравнении Фика): J = — D dC/dx, где J — диффузионный поток [моль∙см-2∙с-1], D — коэффициент диффузии [см2∙с-1], а dC/dx— градиент концентрации.
** Проводимость (g = 1/R) — это характеристика скорости движения любых зарядов через мембрану, а проницаемость (P) отражает, насколько легко частицы могут двигаться через мембрану независимо от того, движутся они или нет. Для ионов эти величины связаны. Например, в фазе быстрой деполяризации при потенциале действия возрастает как проводимость, так и проницаемость мембраны для ионов натрия. Однако зачастую, если проницаемость канала для какого-то иона высока из-за высокого сродства участков поры канала для этого иона, такие ионы будут двигаться через канал медленнее, и проводимость будет ниже, чем для других ионов, для которых канал менее проницаем.
*** Удельная емкость чистого липидного бислоя составляет около 0,8 мкФ/см2, разница между электроемкостью мембраны и липидного бислоя возникает из-за обилия встроенных в мембрану белков.
Библиография
- Hediger M.A. et al. The ABCs of membrane transporters in health and disease (SLC series): Introduction // Mol. Aspects Med. 2013. Vol. 34. P. 95–107.
- Hille B. Ion Channels of Excitable Membranes. Third edit. Sinauer Associates, Inc., 2001.
- Caterina M.J. et al. The capsaicin receptor: A heat-activated ion channel in the pain pathway // Nature. 1997. Vol. 389, № 6653. P. 816–824.
- Jordt S.-E., Tominaga M., Julius D. Acid potentiation of the capsaicin receptor determined by a key extracellular site // Proc. Natl. Acad. Sci. 2000. Vol. 97, № 14. P. 8134–8139.
- Smart D. et al. The endogenous lipid anandamide is a full agonist at the human vanilloid receptor (hVR1) // Br. J. Pharmacol. 2000. Vol. 129, № 2. P. 227–230.
- Nersesyan Y. et al. Oxytocin Modulates Nociception as an Agonist of Pain-Sensing TRPV1 // Cell Rep. 2017. Vol. 21, № 6. P. 1681–1691.
- Berkovic S.F. et al. Human epilepsies: interaction of genetic and acquired factors // Trends Neurosci. 2006. Vol. 29, № 7. P. 391–397.
- Welsh M.J., Smith A.E. Molecular mechanisms of CFTR chloride channel dysfunction in cystic fibrosis // Cell. 1993. Vol. 73, № 7. P. 1251–1254.
- Ruan Y., Liu N., Priori S.G. Sodium channel mutations and arrhythmias // Nat. Rev. Cardiol. 2009. Vol. 6, № 5. P. 337–348.
- Giudicessi J.R., Ackerman M.J. Potassium-channel mutations and cardiac arrhythmias — Diagnosis and therapy // Nat. Rev. Cardiol. Nature Publishing Group, 2012. Vol. 9, № 6. P. 319–332.
- Thomas R.C. Electrogenic sodium pump in nerve and muscle cells // Am. J. Physiol. 1972. Vol. 52, № 3. P. 563–594.
- Morth J.P. et al. Crystal structure of the sodium-potassium pump // Nature. 2007. Vol. 450, № 7172. P. 1043–1049.
- Lodish H. et al. Molecular Cell Biology (5th edition) // Biochemistry and Molecular Biology Education. 2003.
- Scheel O. et al. Voltage-dependent electrogenic chloride/proton exchange by endosomal CLC proteins // Nature. 2005. Vol. 436, № 7049. P. 424–427.
- Artigas P., Gadsby D.C. Ouabain affinity determining residues lie close to the Na/K pump ion pathway. // Proc. Natl. Acad. Sci. U. S. A. 2006. Vol. 103, № 33. P. 12613–12618.
- Li, C., Ramjeesingh, M., Wang, W., Garami, E., Hewryk, M., Lee, D., Rommens, J. M., Galley, K., Bear, C. E. ATPase Activity of the Cystic Fibrosis Transmembrane Conductance Regulator // Journal of Biological Chemistry. 1996. № 45 (271). P. 28463–28468.
- Martineau M. et al. VGLUT1 functions as a glutamate/proton exchanger with chloride channel activity in hippocampal glutamatergic synapses // Nat. Commun. Springer US, 2017. Vol. 8, № 1.
- Ling G., Gerard R.W. The normal membrane potential of frog sartorius fibers // J. Cell. Comp. Physiol. Wiley-Blackwell, 1949. Vol. 34, № 3. P. 383–396.
- Hodgkin A.L., Huxley A.F. Action potentials recorded from inside a nerve fibre // Nature. 1939. Vol. 144. P. 710–711.
- Cole K.S., Curtis H.J. Electric Impedance of the Squid Giant Axon During Activity // J. Gen. Physiol. 1939. Vol. 22, № 5. P. 649–670.
- Cole K.S. Mostly membranes // Annu. Rev. Physiol. 1979. Vol. 41, № 1. P. 1–24.
- Kandel E.R. et al. Principles of Neural Science. Fifth Edit. The McGraw-Hill Companies, Inc., 2013. 1709 p.
- Hodgkin A.L., Huxley A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve // J. Physiol. 1952. Vol. 117. P. 500–544.
- Capes D.L. et al. Domain IV voltage-sensor movement is both sufficient and rate limiting for fast inactivation in sodium channels // J. Gen. Physiol. 2013. Vol. 142, № 2. P. 101–112.
- http://www.physiologyweb.com/lecture_notes/neuronal_action_potential/neuronal_action_potential_na_and_k_concentrations_do_not_change_during_an_action_potential.html
- Bygrave F.L., Benedetti A. What is the concentration of calcium ions in the endoplasmic reticulum? // Cell Calcium. Churchill Livingstone, 1996. Vol. 19, № 6. P. 547–551.
Нашли опечатку? Выделите фрагмент и нажмите Ctrl+Enter.
Биопотенциалы (потенциал действия клеточных мембран, мембранный потенциал покоя)
")
Биопотенциалы (от греческого слова — Bios — жизнь и латинского слова — Potentia — сила) — разность потенциалов между двумя точками живой ткани, которая отражает ее биоэлектрическую активность. Мембраны всех живых клеток в покое находятся в процессе поляризации, то есть имеют разный электрический потенциал внутренней и наружной поверхностей. Разность биопотенциалов между внутренней и наружной поверхностями клеточной мембраны называется мембранным потенциалом покоя. Разность биопотенциалов — величина постоянная и для разных типов клеток возбудимых тканей колеблется от -60 до -100 мВ. Возникновение мембранного потенциала покоя обусловлено разной концентрацией ионов натрия, калия, хлора, кальция снаружи и внутри клетки, а также различной проницаемостью для них мембраны. Так, концентрация ионов калия внутри клетки в 40-50 раз больше, чем в межклеточной жидкости, а концентрация ионов натрия, наоборот, больше снаружи клетки. Та же разница концентраций характерна для ионов кальция. В состоянии покоя мембрана проницаема для ионов калия, так как большинство калиевых каналов открыты, слабопроницаемые для натрия, так как практически все натриевые каналы закрыты, и непроницаемы для органических анионов и хлора, поскольку все каналы для них закрыты. Такое положение мембранных ионных каналов очень важен для генерации мембраннрго потенциала покоя.
Кроме указанных фактов, поляризация клеточной мембраны при открытых калиевых каналах объясняется еще, хотя и небольшим, но существующей утечкой ионов внутриклеточного калия в окружающую среду. Утечка калия создает разницу электрических биопотенциалов в условиях, когда вход ионов натрия в клетку, а также выход из нее органических анионов, которые могли бы нарушить градиент ионов, исключены свойствами клеточной мембраны, в покое.
В указанной выше ситуации на мембране клетки образуется двойной электрический слой: снаружи — катионы, в основном ионы натрия, внутри — анионы, преимущественно органических кислот. Таким образом, выход ионов калия из клетки создает избыток позитивного электрического заряда на внешней поверхности клеточной мембраны, присоединяясь к положительных зарядов ионов натрия. Негативно же заряженные ионы цитоплазмы клеток концентрируются у внутренней поверхности клеточной мембраны, создавая негативный электрический биопотенциал.
При воздействии на клетку раздражителя в ней происходят сложные изменения в обмене веществ, микроструктуре, концентрации ионов и формируется специфическая реакция, обусловленная электрическим биопотенциалом, которая называется потенциалом действия (синоним потенциал возбуждения).
Потенциал действия — очень быстрое колебание мембранного потенциала покоя, формирующее при возбуждении клеток раздражителем пороговой силы. С помощью указанного потенциала действия передаются сигналы от нервов к мышечным клеткам, осуществляется передача информации в нервной системе от одной клетки к другой.
Если достигается критический уровень деполяризации, то происходит быстрое открытие натриевых каналов, которое приводит к лавинообразному поступлению ионов натрия внутрь клетки. Вход ионов натрия в клетку обеспечивает полную деполяризацию клеточной мембраны. В это время поступления позитивных зарядов в клетку вызывает снижение позитивного электрического заряда на внешней стороне мембраны и увеличение его в цитоплазме. Разность биопотенциалов падает до 0, а затем, по мере поступления ионов натрия в клетку, происходит изменение заряда мембранного потенциала покоя.
Внешняя поверхность становится электронегативной по внутренней поверхности клеточной ембраны, то есть происходит инверсия потенциала действия.
Таким образом, вход ионов натрия обеспечивает восходящую фазу пика потенциала действия — деполяризацию. В развитии потенциала действия участвуют, кроме натриевых и калиевых каналов, кальциевые каналы.
Но они способны активизироваться лишь при наличии во внутриклеточной среде факторов, необходимых для реакции фосфорилирования мембранных белков:
- цАМФ
- АТФ
- ионов магния
Ионы кальция играют главную роль в генерации потенциала действия в клетках синусового узла — водителя ритма сердца. В возникновении потенциала действия ионы кальция участвуют путем взноса в процесс деполяризации, осуществляя регуляцию калиевой и натриевой проницаемости клеточных мембран.
Однако ведущая роль кальциевых каналов заключается в обеспечении взаимосвязи между процессами деполяризации клеточной мембраны и процессов внутри клетки. При достижении определенного значения потенциала действия (≈120 мВ) натриевые каналы закрываются, и движение ионов натрия внутрь клетки останавливается (так называемая натриевая инактивация), но продолжается существенный выход ионов калия. Это приводит к приостановке повышения тока действия, пик потенциала действия заканчивается и начинается восстановление поляризации мембраны — реполяризация. В результате натриевой инактивации поток натрия в цитоплазму ослабевает, а увеличение калиевой проницаемости вызывает усиление потока калия в межклеточное пространство. Начинают работать калиевые и натриевые насосы. Сначала натриевый выкачивает ионы натрия наружу, восстанавливая первоначальную разницу концентраций. Затем включается механизм калиевого насоса, который возвращает ионы калия внутрь клетки из межклеточного пространства.
В результате указанных выше процессов внутренняя поверхность клетки вновь приобретает отрицательный заряд по внешней среды. На мембранный потенциал покоя влияет много лекарственных средств, механизм действия которых связан с изменением функциональной активности ионных каналов мембраны. Так, местные анестетики, отдельные противосудорожные препараты (дифенил, карбамазепин) и антиаритмические препараты (этмозин, этацизин, новокаинамид и др.) блокируют натриевые каналы, благодаря чему мешают развитию деполяризации.
Блокаторы медленных кальциевых каналов (верапамил и др.) уменьшают возбудимость клеток синусового узла и таким образом производят противоаритмическое действие. Гипогликемические препараты — производные сульфонилмочевины (глибенкламид и др.) Являются блокаторами АТФ-зависимых калиевых каналов в β-клетках поджелудочной железы. Это способствует деполяризации мембран, открытию потенциалзависимых Са 2+ -каналов, повышению уровня Са 2+ в β-клетках, стимулирует секрецию инсулина. Активаторы К+ -каналов (миноксидил) имеют гипотензивное действие, обусловленное расширением артериол.
^Наверх
Полезно знать
- Барвинок малый (фото, описание, лечебные свойства)
- Ганглий (превертебральный, паравертебральный, вегетативный)
- Бактерии